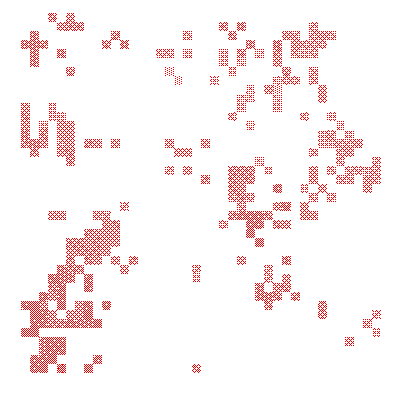
The first three chapters of this thesis deal with evolution in "single" predator-prey systems: a single population of prey interacting with a single population of predators. One of the conclusions is that a stable population dynamical equilibrium of prey and predators may be among the outcomes, but not a priorily. In other cases evolution will lead to populations that persist in a fluctuating way, whereas again in other cases predator or prey will go extinct.
If a single predator-prey system is unstable, interesting things happen when predator and prey are spread over a large region in space. Then, provided individuals move through space relatively slowly, different regions may start to oscillate out of phase. This means that in some regions prey density will be high, while in other regions prey will be virtually absent. A map of the prey population will then show local "patches" of high prey density surrounded by regions devoid of prey; a similar map can be made of the predator population (see Fig. 6 for an example). Such maps will not remain static, of course. Patches of prey will grow and expand. Eventually they will be found and invaded by predators. These predators rapidly multiply, and finally reduce local prey density again. Having depleted the local prey patch, the predators must then search for other patches of prey, that may have been founded in the meantime. This process closely resembles what happens in many mite predator-prey systems.
If predators completely wipe out the prey locally, prey and predator can persist only in a larger whole. Such a dynamic ensemble of local populations that get extinct and become recolonized is called a metapopulation. Metapopulations are of considerable interest, not only for mite predator-prey systems, and have been studied extensively. Should conservation biologists strive for many small nature reserves or just a single big one, when they want to preserve a rare species? The chances that it gets extinct are probably smallest in a big patch, but if it gets extinct it is lost. In an ensemble of patches local extinction will occur more frequently, but empty patches may be recolonized from neighbouring patches.
In a predator-prey metapopulation, natural selection may favour other traits than in a single, "well-mixed" system. In a well-mixed system, natural selection will favour predators that maximize their prey capture rate—because this will result in most offspring. In a metapopulation this may be different. Consider a lucky predator that has just found a new, expanding prey patch. It will start a local population that is exploiting the prey patch. If the predator and its descendants maximize prey capture rate, they should all stay within the prey patch, until they have consumed all prey and have to leave in search of new prey patches. However, there are other strategies strategies to exploit a prey patch. Consider a predator whose descendants have an innate tendency to leave the patch before the prey are extinct. From the viewpoint of the individual, dispersing is a stupid thing to do—after all, why abandon your food if it is uncertain that you will find food elsewhere?—but dispersal helps relatives that stay behind; the local prey population is exploited less heavily and will exist for a longer time. By the time "milkers" have finished the local prey population, the milker foundress will have many more descendants roaming the area than has the "killer" foundress whose descendants all stayed within the patch as long as possible.
The potential advantages of milking are thus very great. But there are disadvantages too: if a milker patch is invaded by a killer, the killer and its descendants quickly consume all prey, spoiling things for the milkers in the patch. It therefore only pays to adopt the milker strategy instead of the killer strategy only if the risk of invasion is rather low. But then, if more predators become killers the risk of invasion decreases—after all, killers do not exploit prey patches in an efficient way and consequently their overall abundance will be low—and the conditions for milkers improve again. Thus there is a balance determining optimal strategies for the exploitation of a prey patch. (This balance is investigated in more detail in Chapter 4.)
If one replaces the terms "prey patch" and "predator" by "host" and "parasite", an interesting analogy arises. A parasite (bacterium, virus or other pathogen) can be said to exploit a host in much the same way as a predator ca be said to exploit a prey patch. When an infective stage enters a host, it multiplies within the host, giving rise to a population of descendants—that is, to a clone. This clone then produces new infective stages to achieve transmission to new hosts. Just as there are different ways to exploit a local prey population, parasites can exploit a host in different ways. "Virulent" parasites are literally "killers"—they opt for quick reproduction to the detriment of their host. "Avirulent" parasites are more of the "milker" type—they reduce ill effects in order to keep their host longer alive. If a parasite clone can monopolize its host, it pays to exploit its host most efficiently—that is, it pays to be avirulent. If a clone has to share its host with other clones, increased virulence is favoured—an attempt to prolong the host"s infective period is ineffective if the other clones do not cooperate. Now if there is a clear case where population dynamics and natural selection are interdependent, it is the case of parasite virulence. When the parasites in the population are avirulent, their efficient host use leads to a high prevalence in the host population. Under population dynamical equilibrium, the "force of infection" is high, which means that the fraction of hosts harbouring multiple parasite clones is also likely to be high; more virulent parasites do better than avirulent ones. However, a change in virulence has population dynamical consequences; more virulent parasites have a lower equilibrium prevalence (less efficient host-use!). This change in population dynamics has, in turn, evolutionary consequences; low parasite prevalence means that the force of infection will be low, which improves conditions for avirulent parasites—they run less risk to lose their host. Thus, the evolution of virulence will be mediated by changes in population dynamics.
Usually, population dynamical models for the interaction between hosts and parasites do not take multiple infection into account. This is a sensible simplification because the arrival of a new parasite individual in a host harbouring a whole population already is unlikely to have many effects. However, for an understanding of the evolution of virulence, multiple infection (or, more precisely, within-host competition among the parasites) should not be ignored. If hosts are infected by single parasite clones, natural selection will favour avirulent ones, whereas if hosts are infected by multiple clones, increased virulence is favoured.