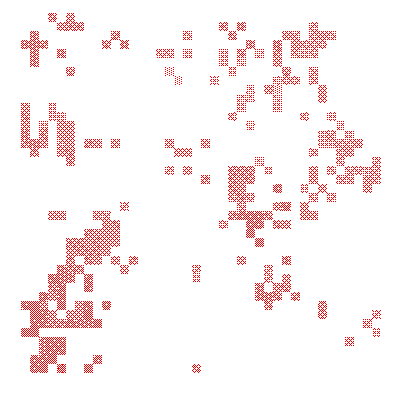
In the 1930s Nicholson and Bailey formulated a simple population dynamical model for predator-prey interactions with non-overlapping generations. Their model turned out to be extremely unstable; an ecological equilibrium of prey and predator exists, but even the slightest perturbation is enough to start a series of increasing oscillations that leads, within a few generations, to the extinction of both prey and predators (Fig. 1). This finding spurred much research, because predator-prey interactions with non-overlapping generations are very common, and somehow these persist. What essential aspect or aspects were overlooked in Nicholson and Bailey"s simple model? Inspired by observation, various modifications of the model were proposed, some of which appeared to promote persistence. But none of these modifications captured the imagination so much as Hassell and May's variant proposed in the 1970s: if the prey population is spread unevenly over space, and if the predators concentrate searching in the areas harbouring most prey, then oscillations would be damped and the ecological equilibrium of prey and predators would be stable (Fig. 2). This model seemed to be very generally applicable. Most prey populations are unevenly distributed; uneven spatial distributions even are the rule rather than the exception. Moreover, predators do preferentially forage in the most profitable patches. Thus, the "aggregative response" of predators to prey density could very well be a very general persistence-promoting mechanism.
Many population ecologists set off to test this hypothesis, by observing spatial distributions of prey, by studying the foraging behaviour of predators, and so forth. A confusing picture emerged, and a debate was started that continues to this day. Of course, uneven prey distributions were found, and predators appeared to have elaborate tactics to search for their prey. What remained a question, however, was whether the strength of the aggregative response would be sufficient to promote persistence. As Hassell & May's model was not designed to describe a specific natural system, but rather to demonstrate the potential consequences of a single factor (spatial distribution) this is not an easy question to decide.
What one can do (as elaborated in Chapter 1 of this thesis) is try to extend Hassell and May's analysis with an evolutionary argument. Hassell and May's model may be a simple "toy world" inhabited by simple organisms, natural selection will operate there all the same. Do spatial population distributions as were assumed by Hassell and May result from searching strategies (of either predator or prey) that are favoured by natural selection?
Consider first evolution of predators only, and take the uneven distribution of the prey as given. Obviously, a single predator should forage in the patch containing most prey. However, if a large number of other predators will do the same, natural selection may actually favour predators that escape the crowd and forage in low-prey-density patches! Thus, natural selection will put an upper bound to the strength of the "aggregative response", and this means that the scope for persistence is reduced.
When not only the predators evolve but also the prey, the situation changes profoundly. Prey will do best by avoiding patches with a high numbers of conspecifics, because most predators will be found there. An illustration is given in Figure 3 (which is the same as Fig. 5 from Chapter 1). It depicts a hypothetical environment consisting of five patches of food for the prey, where one of the patches is of much better quality (Fig. 3a). In absence of predators, the prey will obviously do best by moving to patch number one. But if all prey move into a single patch, the predators will all follow. Then the better quality of the patch may completely offset by the risk of predation, which means that a mutant prey that selects one of the low-quality patches will get more offspring. The prey strategy to select patch one is therefore not evolutionarily stable. But then, if enough prey seek refuge in the low-quality patches, a mutant predator that selects one of the a low-quality patches can invade. Eventually evolution will lead to a situation in which it pays for no individual to prefer any one patch over another. This situation is depicted in Figure 3b. It can be seen that for uneven spatial distributions (light shading: distribution of the prey; dark shading: of the predators) a large difference in patch quality must be assumed. As ecological stability requires population distributions roughly of the type as shown in Figure 3b, the conditions for ecological stability are quite narrow.
If the population dynamical equilibrium of predator and prey is stable, predator and prey are likely to persist—obviously. However, this does not mean that an unstable population dynamical equilibrium will always lead to extinction. Indeed, population dynamical analysis has revealed that in many cases populations remain fluctuating but nevertheless do not go extinct even in the long run. Such fluctuations may be regular (which is called a limit cycle, see Fig. 4) or irregular (in which case the dynamics are called chaotic, see Fig. 5).
In Chapter 1 we concluded that the conditions for a stable ecological equilibrium of prey and predator were quite narrow. But we found that the conditions for non-equilibrium persistence—in the form cyclic or chaotic fluctuations—may occur in a much wider range of conditions (Chapter CH2). This holds especially when predators and prey can flexibly react to their surroundings—that is, seek out the best patches depending on the circumstances. This conclusion would suggest that the refined abilities of many prey and predators to assess and react to their environment may have far-reaching consequences; the availability of information will then be an important factor determining population dynamics.